|
Một số điểm mới trong nghiên cứu miễn dịch bệnh nhiễm vi rút, vi khuẩn và ký sinh trùng
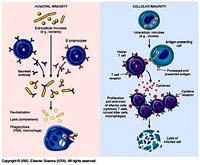
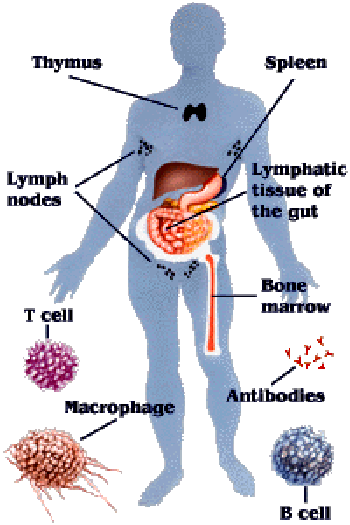
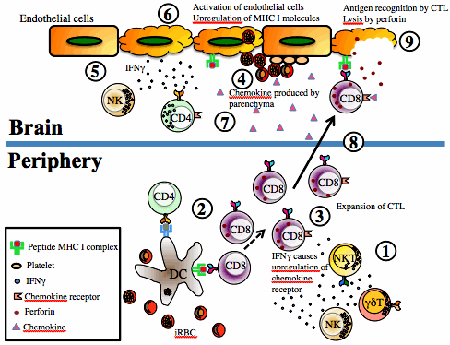
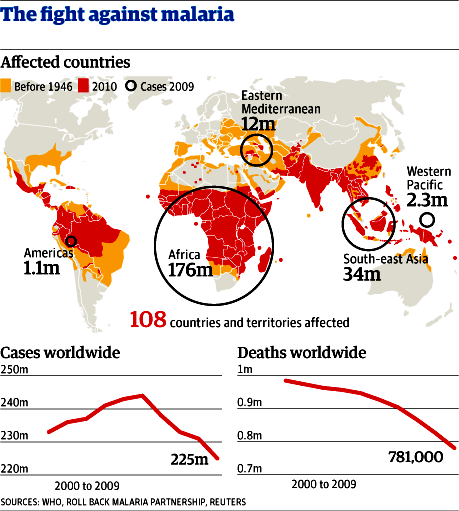
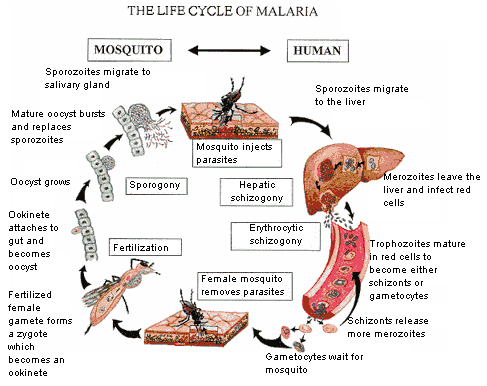
Đáp ứng miễn dịch chống một số bệnh nhiễm virus, vi khuẩn và ký sinh trùng; Ký sinh trùng sốt rét “lẩn tránh” hệ miễn dịch; Ẩn mình vào trong tế bào hình cây của lách: Vẫn còn nhiều vấn đề chưa biết đến!; Miễn dịch dịch thể đặc hiệu từng giai đoạn phát triển KSTSR; Cho muỗi đốt để miễn dịch với bệnh sốt rét - Nên chăng? Đáp ứng miễn dịch chống một số bệnh nhiễm virus, vi khuẩn và ký sinh trùng Để cho một vi sinh vật gây ra được nhiễm trùng cho một túc chủ nhậy cảm thì cần phải có các sự kiện phối hợp với nhau làm ngăn cản sức đề kháng đặc hiệu và không đặc hiệu của túc chủ đó. Nhìn chung các vi sinh vật có nhiều cách để thoát khỏi sự tấn công miễn dịch của cơ thể túc chủ. Nhiều vi sinh vật giảm tính kháng nguyên của chúng, hoặc cư trú bên trong tế bào của túc chủ để’n tránh’sự tấn công miễn dịch, hoặc bằng cách làm trụi các kháng nguyên màng của chúng. Các vi sinh vật bắt chước các phân tử màng của tế bào túc chủ hoặc bằng cách biểu hiện các phân tử có cấu trúc tương tự như các phân tử của màng tế bào túc chủ, hoặc bằng cách thâu nạp các phân tử của túc chủ để’ngụy trang’cho mình. Trong một số trường hợp vi sinh vật có khả năng gây ức chế một cách chọn lọc các đáp ứng miễn dịch hoặc điều biến các đáp ứng miễn dịch để làm chệch hướng miễn dịch (tức là sinh ra một đáp ứng miễn dịch không nguy hại gì đến chúng). Sự thay đổi về cấu trúc các kháng nguyên bề mặt cũng là một cách để các vi sinh vật thoát khỏi tác dụng của hệ thống miễn dịch. Trong chương này chúng ta sẽ thảo luận những quan niệm liên quan tới miễn dịch chống lại các virus, vi khuẩn, các đơn bào, giun sán - Đó là những vi sinh vật chính gây nên các bệnh nhiễm trùng ở người. Đáp ứng miễn dịch trong nhiễm virus Cơ thể có một số cơ chế miễn dịch đặc hiệu cùng với các cơ chế đề kháng không đặc hiệu nhằm mục đích loại bỏ các virus lây nhiễm. Đồng thời virus cũng hoạt động để vượt qua một hoặc nhiều cơ chế đề kháng này để kéo dài sự tồn tại của chúng. Những biểu hiện của nhiễm trùng sẽ phụ thuộc vào cách mà các cơ chế đề kháng của túc chủ chống lại một cách có hiệu quả các’mánh kho’của virus. Trung hoà virus bằng các kháng thể Các kháng thể đặc hiệu với các kháng nguyên bề mặt của virus thường đóng vai trò quyết định trong việc hạn chế sự lan tràn của virus khi nhiễm trùng cấp tính và ngăn cản tái nhiễm. Phần lớn các virus có các phân tử thụ thể bề mặt có khả năng khởi động quá trình nhiễm trùng bằng cách gắn một cách đặc hiệu với các phân tử màng của tế bào túc chủ. Ví dụ virus cúm kết hợp với các gốc acid sialic có trong các glycoprotein và glycolipid của màng tế bào; virus rhino kết hợp với các phân tử kết dính (ICAM); virus Epstein - Barr kết hợp với các thụ thể type II dành cho bổ thể trên bề mặt tế bào B. Nếu các kháng thể được sinh ra để chống lại các thụ thể của virus thì chúng có thể phong bế sự nhiễm trùng bằng cách ngăn cản sự kết hợp của các hạt virus vào tế bào túc chủ. IgA tiết trong các dịch tiết của các màng nhầy có một vai trò quan trọng trong sức đề kháng của túc chủ chống lại virus bằng cách ngăn cản sự gắn của virus vào các tế bào biểu mô của niêm mạc. Ưu điểm của vaccine bại liệt giảm độc lực uống chính là ở chỗ vaccine này kích thích sự sản xuất của IgA tiết có tác dụng ngăn cản sự kết hợp của virus bại liệt với các tế bào niêm mạc trong đường tiêu hóa. Các kháng thể còn có thể trung hòa virus bằng các cách khác xẩy ra sau khi virus đã bám vào các tế bào túc chủ. Trong một số trường hợp các kháng thể có thể phong bế sự thâm nhập của virus vào tế bào bằng cách kết hợp với các quyết định kháng nguyên cần thiết cho việc liên hợp của vỏ virus với màng bào tương. Nếu các kháng thể được hình thành là loại hoạt hóa bổ thể thì chúng có thể phá hủy vỏ của virus. Các kháng thể cũng có thể làm ngưng kết các hạt virus và hoạt động như một tác nhân opsonin hóa thúc đẩy hiện tượng thực bào các hạt virus. Các cơ chế miễn dịch tế bào chống virus Mặc dù kháng thể có một vai trò quan trọng trong việc ngăn cản sự lan tràn của virus ở giai đoạn nhiễm trùng cấp, nhưng chúng thường không có khả năng loại bỏ được virus khi nhiễm trùng đã xuất hiện, đặc biệt khi virus có khả năng gây ra trạng thái ẩn (AND của chúng được cài cắm vào AND nhiễm sắc thể của tế bào túc chủ). Khi nhiễm trùng đã xẩy ra thì các cơ chế đáp ứng miễn dịch qua trung gian tế bào chiếm vị trí quan trọng nhất trong sức đề kháng của túc chủ. Các tế bào T H hoạt hóa sản sinh ra một loạt cytokine hoạt động trực tiếp hoặc gián tiếp chống virus. IFN - g hoạt động một cách trực tiếp bằng cách sinh ra trạng thái chống virus bên trong tế bào. Chúng cũng có thể có hoạt tính kháng virus gián tiếp thông qua việc kích thích sinh IL - 2 và IFN - g có tác dụng hoạt hóa tế bào NK. Những tế bào này đóng vai trò quan trọng trong sức đề kháng của túc chủ trong những ngày đầu mới nhiễm của nhiều loại virus khi đáp ứng miễn dịch qua trung gian tế bào đặc hiệu chưa hình thành. Trong phần lớn các trường hợp nhiễm virus hiện tượng gây độc tế bào bởi lympho TCđặc hiệu sẽ xuất hiện 3 - 4 ngày sau nhiễm virus và đạt điểm cực đại sau 1 tuần rồi sau đó giảm xuống. Hiện tượng gây độc tế bào bởi tế bào T Cđặc hiệu sẽ tiêu diệt các tế bào đã nhiễm virus và vì vậy loại bỏ nguồn sản sinh virus mới. Có thể chứng minh vai trò của các tế bào T C trong việc đề kháng chống virus bằng cách gây miễn dịch vay mượn (chuyển các tế bào T C từ một cơ thể đã nhiễm virus sang một cơ thể bình thường thì cơ thể này có khả năng ngăn cản được sự nhiễm virus đó). Virus né tránh các cơ chế đề kháng của túc chủ Một số virus có thể thoát khỏi sức tấn công miễn dịch bằng cách thay đổi kháng nguyên của chúng. Trong trường hợp nhiễm virus cúm, sự thay đổi kháng nguyên liên tục đãẫn đến sự hình thành thường xuyên các chủng virus gây bệnh mới. Các hạt virus cúm có dạng hình cầu hoặc hình ô - val sù sì với đường kính trung bình là 90 - 100 nm, được bao bọc xung quanh bởi một lớp vỏ bên ngoài đó là một màng lipid kép lấy được từ màng bào tương của tế bào túc chủ đã bị nhiễm virus này trong quá trình thâm nhập. Có 2 glycoprotein lớp vỏ này đó là hemagglutinin (HA) và neuramini ase (NA) hình thành các gai nhô ra mà có thể trông thấy được dưới kính hiển vi điện tử. Các gai HA trong dạng trimer chịu trách nhiệm gắn virus vào tế bào túc chủ. Mỗi hạt virus có khoảng 1.000 gai HA. Trimer HA kết hợp với nhóm acid sialic có trên phân tử glycoprotein hoặc glycolypid của tế bào túc chủ. Neuraminidase là một enzyme có thể phân cắt acid N - acetylneuramic khỏi phân tử glycoprotein của virus hoặc phân tử glycoprotein màng tế bào túc chủ, thúc đẩy sự lan tràn của virus từ các tế bào túc chủ đã bị nhiễm. Trong vỏ còn có một lớp protein đáy trơ bao quanh vỏ nhân trong đó có chứa 8 sợi ARN xoắn đơn gắn với protein và ARN polymerase (hình 1). Mỗi sợi ARN sẽ mã hóa một protein khác nhau của virus cúm khác nhau. Có 3 type cơ bản của virus cúm (A, B, C) phân biệt bởi sự khác nhau về nucleoprotein và các protein đáy. Type A là phổ biến nhất và thường gây nên những đại dịch cúm ở người. Sự thay đổi kháng nguyên trong HA và NA lại cho phép phân type A thành các phân type nhỏ theo thuật ngữ của Tổ chức Y tế Thế giới: mỗi chủng virus được xác định bởi nguồn gốc túc chủ của nó (nếu phông phải là người), nguồn gốc địa lý, số chủng, năm phân lập được và loại kháng nguyên HA và NA. Ví dụ A/ SW/ Iowa/ 15/ 30 (H1N1) là tên hiệu của chủng số 15 phân lập được từ cừu ở Iowa vào năm 1930; A/ Hongkong/ 1/ 68/ (H3N2) là tên chủng số 1 phân lập được ở người tại Hồng Kông vào năm 1968. Hai chủng trên có các kháng nguyên H và N khác nhau. Nét đặc trưng của virus cúm là sự thay đổi kháng nguyên của chúng. Virus có thể thay đổi kháng nguyên bề mặt một cách hoàn toàn đến nỗi đáp ứng miễn dịch đối với virus trong một vụ dịch sẽ không còn tác dụng chống lại virus trong vụ dịch trước đó. Sự thay đổi kháng nguyên chủ yếu xẩy ra do sự thay đổi các gai HA và NA nhô ra từ vỏ virus (hình 2). Có hai cơ chế khác nhau làm thay đổi kháng nguyên HA và NA đó là’cải biên’kháng nguyên (antigenic rift) và’thay mới’kháng nguyên (antigenic shift). Cải biên kháng nguyên bao gồm một loạt các biến dị điểm ngẫu nhiên xuất hiện một cách trình tự dẫn đến những thay đổi nhỏ trong HA và NA. Sự thay mới kháng nguyên dẫn đến hình thành đột nhiên một type mới của virus cúm có HA và NA hoàn toàn khác biệt với virus trước đó. Virus cúm ở người lần đầu tiên được phân lập vào năm 1934 với ký hiệu là H0N1. Type này tồn tại tiềm tàng đến năm 1947, lúc này có một sự thay mới kháng nguyên sinh ra một type mới với ký hiệu là H1N1. Type H1N1 thay thế type trước và lan tràn trên thế giới đến năm 1957 thì xuất hiện type H2N2. Type H2N2 lưu hành trên thế giới trong suốt thập kỷ 60 và đến năm 1968 biến đổi thành type H3N2. Sự thay mới kháng nguyên xuất hiện gần đây nhất xẩy ra vào năm 1977 làm tái xuất hiện type H1N1. Trong mỗi lần thay đổi kháng nguyên đều xẩy ra sự thay đổi trình tự các acid amine căn bản trong cấu trúc HA và NA dẫn đến thay đổi kháng nguyên rõ rệt mà hệ thống miễn dịch của túc chủ chưa có trí nhớ miễn dịch đối với kháng nguyên này. Như vậy thay mới kháng nguyên xẩy ra ở một quần thể chưa có chuẩn bị về miễn dịch làm xuất hiện các đại dịch cúm cho loài người như đã xẩy ra. Giữa những vụ đại dịch virus cúm vẫn có sự cải biên kháng nguyên gây ra những thay đổi không nhiều, đáp ứng miễn dịch vẫn xẩy ra để chống lại các chủng virus cúm này. Khi một cá thể đã bị nhiễm 1 chủng virus cúm nhất định và sinh ra một đáp ứng miễn dịch thì chủng virus tương tự chủng này sẽ bị loại bỏ. Tuy nhiên sự tích lũy các biến dị điểm cũng làm thay đổi tính kháng nguyên của một số chủng đủ để chúng có thể thoát khỏi sự loại bỏ do miễn dịch. Những chủng này là những chủng mới, gây ra một chu kỳ dịch địa phương khác. Vai trò của kháng thể trong việc lựa chọn miễn dịch như vậy có thể được chứng minh trong phòng thí nghiệm bằng cách trộn một chủng virus cúm với kháng thể đơn clone đặc hiệu cho chủng này và sau đó cấy virus trong tế bào. Kháng thể trung hòa tất cả các hạt virus không thay đổi, chỉ có các hạt virus xẩy ra những biến dị dẫn đến thay đổi tính kháng nguyên sẽ không bị kháng thể trung hòa và nhân lên. Sau một thời gian ngắn có thể xác định được chủng virus cúm mới này. Đảo lộn kháng nguyên được giả thiết là do sự tái liên kết giữa các virus cúm của người với các virus cúm từ động vật khác nhau bao gồm ngựa, lợn và vịt. Thật vậy, virus cúm chứa 8 sợi ARN xoắn đơn tách biệt nhau, điều này tạo ra khả năng tái liên kết của các sợi ARN của các hạt virus của người với các sợi ARN của các hạt virus động vật. Nếu như một tế bào riêng lẻ bị nhiễm đồng thời cả hai loại virus. Năm 1971 Webster. R. G và Campbell. C. H đã nêu lên những bằng chứng về sự tái liên kết di truyền in vivo của virus cúm A của người và của lợn nhà. Sau khi gây nhiễm cho 1 lợn đồng thời virus Hồng Kông của người (H3N2) và virus cừu (H1N1) thì người ta đã phát hiện được các hạt virus có H3N1. Trong một số trường hợp sự thay mới kháng nguyên rõ rệt đã làm tái xuất hiện một chủng virus mà trước đây đã tồn tại ẩn nấp trong nhiều thập kỷ. Ví dụ tháng 5 năm 1977 một chủng virus cúm A/uSSR/ 77 (H1N1) đã được chứng minh là giống với một chủng gây nên vụ dịch 27 năm về trước. Trong trạng thái đóng băng thì virus có thể tồn tại nhiều năm, khi các virus này được tái xuất hiện thì các HA và NA không phải là hoàn toàn mới. Tuy nhiên chúng sẽ được nhận biết bởi hệ thống miễn dịch như là chủng mới bởi vì không có các tế bào mang trí nhớ miễn dịch đặc hiệu cho các kháng nguyên của chủng virus này. Vì vậy, trên quan điểm miễn dịch học thì sự tái xuất hiện của một chủng virus cúm A trước đây có thể có những hiệu quả tương tự như một sự thay mới kháng nguyên sinh ra một type mới. Sự thay đổi kháng nguyên trong virus Rhino làm cho người ta không thể sản xuất được các vaccine hữu hiệu. Không đâu có sự thay đổi kháng nguyên lớn hơn sự thay đổi kháng nguyên ở virus HIV. Người ta dự đoán rằng HIV có biến dị 65 lần lớn hơn virus cúm. Một số lớn virus nétránh đáp ứng miễn dịch bằng cách sinh ra ức chế miễn dịch. Trong số này có paramyxovirus gây bệnh quai bị, virus sởi, virus EpsteinBarr, virus cự bào (cytomegalovirus) và HIV. Trong một số trường hợp tình trạng ức chế miễn dịch xẩy ra là do nhiễm virus trực tiếp và các lympho bào và đại thực bào, do vậy virus có thể phá hủy trực tiếp các tế bào miễn dịch bằng các cơ chế làm tan tế bào hoặc làm thay đổi chức năng của các tế bào này. Trong các trường hợp khác, ức chế miễn dịch xuất hiện do sự mất cân bằng cytokine. Ví dụ gene virus Epstein - Barr tương tự với gene IL - 10, mà IL - 10 có tác dụng ức chế tế bào TH1 sản xuất cytokine vì vậy dẫn đến làm giảm IL - 2 và IFN - g. Đáp ứng miễn dịch trong nhiễm vi khuẩn Miễn dịch trong nhiễm các vi khuẩn đạt được chủ yếu nhờ các kháng thể, trừ khi vi khuẩn có khả năng mọc được bên trong tế bào. Đối với những trường hợp vi khuẩn sống bên trong tế bào thì đáp ứng miễn dịch qua trung gian tế bào lại có vai trò quan trọng. Vi khuẩn xâm nhập vào cơ thể hoặc là theo các con đường tự nhiên (đường hô hấp, tiêu hóa, tiết niệu sinh dục) hoặc qua các con đường không tự nhiên (qua các vết thương của da và niêm mạc). Cơ thể túc chủ sẽ sinh ra đáp ứng miễn dịch với những mức độ khác nhau, một phần phụ thuộc vào số lượng vi khuẩn xâm nhập ít và độc lực của chúng thấp thì các tế bào làm nhiệm vụ thực bào tại chỗ có thể đảm đương được việc đề kháng không đặc hiệu và loại bỏ được các vi khuẩn này. Nếu số lượng vi khuẩn lớn hơn và có độc lực cao hơn thì sẽ kích thích cơ thể sinh ra đáp ứng miễn dịch. Đáp ứng miễn dịch chống lại các vi khuẩn ký sinh bên ngoài và bên trong tế bào. Những vi khuẩn ký sinh bên ngoài (vi khuẩn ngoại bào) tế bào sẽ kích thích cơ thể sinh ra kháng thể, kháng thể này thường được tiết bởi các tế bào plasma trong các hạch lympho khu vực hoặc trong lớp dưới niêm mạc của đường hô hấp hoặc đường tiêu hóa. Bằng một số cách, kháng thể sẽ phá hủy các vi khuẩn. Những kháng thể gắn với những kháng nguyên lộ diện trên bề mặt của vi khuẩn cùng với thành phần C3b của bổ thể sẽ họat động như một yếu tố opsonin làm tăng hiện tượng thực bào. Sự hoạt hóa hệ thống bổ thể do kháng thể (con đường cổ điển) làm xuất hiện một số phân tử có tác dụng phát triển và khuyếch đại đáp ứng viêm. Ví dụ khi bổ thể bị cắt thành các sản phẩm C3a, C4a, C5a chúng sẽ hoạt động như các độc tố gây phản vệ làm cho các tế bào mast gây tăng tính thấm thành mạch tạo điều kiện thuận lợi cho các lympho bào và bạch cầu trung tính thoát mạch tiến đến ổ viêm. Các sản phẩm khác của sự phân cắt bổ thể có tác dụng như các yếu tố hóa hướng động đối với bạch cầu trung tính và vì vậy sẽ làm tăng số lượng các tế bào làm nhiệm vụ thực bào tại nơi nhiễm trùng. Đối với một số vi khuẩn, chủ yếu là các vi khuẩn gram âm, sự hoạt hóa bổ thể có thể dẫn đến dung giải vi khuẩn. Nếu vi khuẩn tiết ngoại độc tố hoặc nội độc tố thì kháng thể có thể kết hợp với độc tố và trung hòa chúng. Phức hợp kháng thể - Độc tố có thể được thanh lọc bởi các tế bào làm nhiệm vụ thực bào. Các trường hợp nhiễm vi khuẩn ký sinh bên trong tế bào (vi khuẩn nội bào), đặc biệt là ký sinh trong các tế bào thực bào, sẽ kích thích cơ thể sinh đáp ứng miễn dịch qua trung gian tế bào. Trong đáp ứng này các tế bào TDTH sẽ tiết ra các cytokine, đặc biệt là IFN - (, có tác dụng hoạt hóa đại thực bào làm tăng khả năng nuốt và giết các vi khuẩn ký sinh bên trong tế bào. Vi khuẩn né tránh các cơ chế đề kháng của túc chủ Vi khuẩn có các cơ chế khác nhau để tạo điều kiện cho chúng hình thành các khuẩn lạc trên niêm mạc của túc chủ và thoát khỏi các đáp ứng miễn dịch của túc chủ (bảng 1). Một số vi khuẩn gram âm có các pili (là những vi lông roi dài) làm cho chúng có khả năng bám vào niêm mạc ruột hoặc niêm mạc đường tiết niệu sinh dục, đó là bước đầu tiên của quá trình nhiễm. Các vi khuẩn như Bordetella pertussis (gây ho gà) tiết ra các phân tử kết dính để gắn vào vi khuẩn khác hoặc các tế bào biểu mô trụ của đường hô hấp trên. IgA tiết có khả năng ngăn cản vi khuẩn bám vào các tế bào biểu mô. Tuy nhiên một số vi khuẩn như Neisseria gonorrhoeae, Hemophilus influenzae và Neisseria meningitidis tiết ra các protease có tác dụng phân cắt IgA tiết ở vùng bản lề. Mảnh Fab và mảnh Fc khi được tách ra từ phân tử IgA tiết nguyên vẹn có đời sống bán hủy ngắn và không còn khả năng gây ngưng kết vi khuẩn. Các vi khuẩn còn có thể thoát khỏi tác dụng của IgA tiết và vì vậy tăng khả năng gắn vào tế bào biểu mô bằng cách thay đổi kháng nguyên bề mặt của chúng, ví dụ như N. Gonorrhoeae có khả năng gắn vào tế bào biểu mô của niệu đạo hoặc cổ tử cung do chúng có các pili. Vi khuẩn này có chứa một thành phần protein của pili được gọi là pilin có cấu trúc của vùng hằng định, vùng thay đổi và vùng các acid amin siêu biến. Sự thay đổi thứ tự các acid amin của pilin diễn ra do sự bố trí lại của các gene mã hóa protein này. Locus pilin có chứa 1 hoặc 2 gene biểu hiện và 10 - 20 gene im lặng. Mỗi 1 gene được phân bố thành 6 vùng được gọi là các minicassette. Sự thay đổi pilin xẩy ra trong quá trình biến trạng gene trong đó một hoặc nhiều minicassette từ trạng thái gene im lặng sẽ thay thế một minicassette của gene biểu hiện (hình 4). Quá trình này sinh ra tính đa dạng kháng nguyên của protein pilin tương tự như sự tái tổ hợp gene trong quá trình sản xuất globulin miễn dịch. Sự thay đổi liên tục trong cấu trúc pilin có thể ảnh hưởng đến khả năng gây bệnh của N. Gonorrhoeae bằng cách làm tăng sự biểu hiện của pili để bám chắc vào tế bào biểu mô. Ngoài ra sự thay đổi liên tục của pilin cho phép vi khuẩn thoát khỏi sự trung hòa bởi kháng thể. Nhiều vi khuẩn có các cách đề kháng với hiện tượng thực bào hoặc thoát khỏi các đáp ứng miễn dịch có bổ thể tham gia. Ví dụ một số vi khuẩn có cấu trúc bề mặt mà cấu trúc này có thể ức chế hiện tượng thực bào. Streptococus pneumoniae có cấu trúc vách là polysaccarit rất hiệu quả trong việc ngăn cản hiện tượng thực bào. Streptococus pyogenes có một protein bề mặt gọi là protein M cũng ức chế hiện tượng thực bào. Các Straphylococci gây bệnh tiết ra một enzyme coagulase xúc tác để tạo thành màng fibrin bao quanh vi khuẩn, nhờ vậy thoát khỏi các tế bào thực bào. Một số vi khuẩn có tác dụng trên hệ thống bổ thể. Ví dụ các vi khuẩn gram âm có các chuỗi bên dài trên nửa lipid A của polisaccharit lõi trong thành tế bào, các chuỗi này có tác dụng kháng lại hiện tượng dung giải tế bào do bổ thể gây nên. Pseudomonas tiết ra elastase có tác dụng bất hoạt cả với C3a và C5a, vì vậy làm giảm phản ứng viêm tại chỗ. Giai đoạn đáp ứng của túc chủ -Bám vào tế bào IgA tiết ngăn cản túc chủ vi khuẩn bám vào tế bào; -Cơ chế né tránh của vi khuẩn; -Tiết các protease phân cắt IgA dimer (Neisseria meningitidis, N. Gonorrhoeae, Haemophilus Influenzae); -Thay đổi kháng nguyên ở vị trí bám (thay đổi cấu trúc các pili của N. gonorrhoeae); -Nhân lên, xâm nhập vào mô. Gây tổn thương tế bào túc chủ bằng độc tố Một số vi khuẩn thoát khỏi các cơ chế đề kháng của túc chủ bởi khả năng của chúng sống bên trong các tế bào thực bào. M. tuberculosis và M. leprae có thể thoát ra khỏi phagolysosom và nhân lên trong môi trường bào tương. Những vi khuẩn khác như M. avium và Chlamy ia phong bế sự liên hợp của lysosom với phagolysosom. Một số vi khuẩn có khả năng kháng lại tác dụng của các gốc tự do sinh ra trong quá trình thực bào; -Sự tham gia của đáp ứng miễn dịch vào bệnh sinh của các bệnh do vi khuẩn Nhiều trường hợp bệnh gây ra không phải do vi khuẩn mà lại là do đáp ứng miễn dịch chống vi khuẩn. Trong một số trường hợp nhiễm vi khuẩn gram âm, các nội độc tố bản chất là lypopolysaccarit hoạt hóa đại thực bào làm giải phóng nhiều IL - 1 và TNF(Các cytokine này tham gia vào cơ chế sốc do nhiễm khuẩn huyết; -Trong các trường hợp nhiễm độc thức ăn do Staphylococus và hội chứng sốc do nhiễm độc lượng cytokine cao trong máu cũng gây ra những triệu chứng nhất định. Khi nhiễm độc thức ăn do Staphylococus a ureus cơ thể bị nhiễm độc do độc tố ruột của Staphylococus a ureus (kí hiệu của các độc tố ruột này là Ses), có thể phân loại độc tố ruột thành 5 nhóm khác nhau A, B, C, D, E; -Các Thực bào (opsonin hóa bởi C3b hoặc bởi kháng thể. Tan vi khuẩn bởi bổ thể và đáp ứng viêm tại chỗ -Ngưng tập bởi kháng thể; -Trung hòa độc tố bằng kháng thểTạo ra các cấu trúc trên bề mặt (ví dụ như tạo ra nang polysaccharide, protein M, vỏ fibrin) có tác dụng ức chế các tế bào thực bào; -Phát triển các cơ chế để có thể sống được bên trong các tế bào thực bào; -Làm cho các đại thực bào chết do apoptosis (chết tế bào theo chương trình) ví dụ như Shigella flexneri; -Các vi khuẩn gramương phát triển các cơ chế kháng lại sự dung giải bởi bổ thể; -Một số vi khuẩn gram âm cài chuỗi bên dài trong lipopolysaccharide vách của vi khuẩn vào phức hợp tấn công màng để’lấp’lỗ thủng do phức hợp tấn công màng tạo ra; -Tiết elastase làm bất hoạt C3a và C5a (Pseudomonas); -Tiết hyaluroni ase tăng cường sự xâm nhập của vi khuẩn độc tố ruột này hoạt động như những chất kích thích phân bào hoạt hóa tất cả các tế bào T biểu hiện một họ gene V (của thụ thể tế bào T. Vì vậy, những độc tố ruột này được gọi là các’siêu kháng nguyên’. Các cytokine do các tế bào Th giải phóng ra khi được hoạt hóa bởi các siêu kháng nguyên này sẽ gây nên nhiều triệu chứng như sốt, ỉa chẩy, sốc trong nhiễm độc thức ăn do Staphylococus aureus. Các triệu chứng như trên cũng xuất hiện trong hội chứng sốc nhiễm độc, một bệnh có thể gây chết người. Trong trường hợp này ngoại độc tố của tụ cầu được gọi là độc tố 1 gây hội chứng sốc, hoạt động như một siêu kháng nguyên kích thích các tế bào Th hoạt hóa đại thực bào tiết ra nhiều TNF. Các vi khuẩn sống bên trong tế bào thực bào có khả năng hoạt hóa tế bào TDTH dẫn tới phá hủy các mô theo cơ chế quá mẫn muộn. Các cytokine do các tế bào TDTH hoạt hóa tiết ra sẽ chiêu mộvà tập trung các đại thực bào rồi hoạt hóa chúng để hình thành các u hạt. Các enzyme lysosom được giải phóng từ các u hạt này sẽ gây ra hoại tử đáng kể các mô. Ví dụ điển hình là trường hợp loét trong quá mẫn muộn đối với M. tuberculosis. Đáp ứng miễn dịch trong nhiễm các đơn bào. Các đơn bào gây ra nhiều bệnh nguy hiểm cho người như bệnh do amip, bệnh Chaga, bệnh ngủ Châu Phi, sốt rét, bệnh do leishmania, bệnh do toxoplasma. Sự phát triển của đáp ứng miễn dịch và hiệu quả của chúng phụ thuộc một phần vào nơi cư trú của ký sinh trùng trong cơ thể túc chủ. Nhiều loại đơn bào có giai đoạn lưu hành trong òng máu, trong giai đoạn này thì kháng thể là có hiệu quả với chúng nhất. Nhiều loại đơn bào có khả năng sống bên trong tế bào thì chỉ có đáp ứng miễn dịch qua trung gian tế bào mới có hiệu qủa đối với chúng; Đáp ứng của túc chủ với Plasmodium spp. Trong những vùng dịch tễ sốt rét đáp ứng miễn dịch với ký sinh trùng sốt rét thường yếu. Những trẻ dưới 14 tuổi thường có đáp ứng miễn dịch thấp và hay bị bệnh nhất. Trong một số vùng tỷ lệ tử vong ở trẻ em do sốt rét lên tới 50% và hàng năm trên thế giới có tới 1 triệu trẻ chết vì sốt rét. Đáp ứng miễn dịch thấp với ký sinh trùng sốt rét được thể hiện bằng nồng độ kháng thể thấp đối với giai đoạn thoa trùng. Chỉ có 22% số trẻ sống trong vùng dịch tễ sốt rét có kháng thể kháng thoa trùng, trong khi đó 84% người lớn có kháng thể này. Mặc dù vậy ở người lớn mức độ miễn dịch cũng thấp, tuy nhiên phần lớn người sống trong vùng dịch tễ đã bị nhiễm ít ký sinh trùng trong một thời gian dài. Có nhiều yếu tố làm cho đáp ứng miễn dịch chống Plasmodium thấp. Những thay đổi từ thoa trùng thành thành thể phân liệt và giao bào đã làm cho ký sinh trùng thay đổi kháng nguyên bề mặt của chúng. Các giai đoạn sống bên trong tế bào gan và hồng cầu làm giảm mức độ đáp ứng miễn dịch và làm cho chúng có thể nhân lên do tránh được sự tấn công của hệ thống miễn dịch. Hơn thế nữa, thoa trùng tuần hoàn trong máu chỉ 30 phút trước khi chui vào tế bào gan vì vậy sự hoạt hóa miễn dịch khó có thể xuất hiện trong một thời gian ngắn như vậy. Người ta còn phát hiện thấy thoa trùng của Plasmodium được bao phủ bởi 1 protein có trọng lượng phân tử 45 kDa được gọi là kháng nguyên bao quanh thoa trùng (circumsporozoit antigen - CS) cho nên ngay cả khi kháng thể chống thoa trùng xuất hiện thì Plasmodium vẫn thoát khỏi đáp ứng này bằng cách bong các kháng nguyên CS che phủ trên bề mặt. Vaccine chống sốt rét. Một điều rõ ràng là vaccine chống sốt rét muốn có hiệu quả thì cần phải kích thích tối đa các cơ chế đề kháng miễn dịch bảo vệ. Nhưng tiếc thay chúng ta còn biết rất ít về vai trò của đáp ứng miễn dịch thể dịch cũng như đáp ứng miễn dịch qua trung gian tế bào trong việc bảo vệ cơ thể chống lại bệnh sốt rét. Phần lớn những tiếp cận về vaccine chống sốt rét tập trung vào giai đoạn thoa trùng. Người ta đã chế thử một vaccine thử nghiệm gồm các thoa trùng Plasmodium giảm độc lực bằng chiếu tia X. Vaccine này đã được thử cho chuột nhắt, khỉ, người tình nguyện và thấy rằng có cả đáp ứng miễn dịch thể dịch lẫn đáp ứng miễn dịch qua trung gian tế bào chống lại thoa trùng. Các chuột nhắt được miễn dịch sẽ được bảo vệ khi thử thách lại với nhiễm Plasmodium của chuột. Kết quả không như mong đợi, vaccine thoa trùng chiếy tia X không phải là một cách tiếp cận cho việc miễn dịch hàng triệu người sống trong vùng dịch tễ, vì người ta dự tính rằng cần phải có một cơ sở rất lớn nuôi muỗi mới có đủ thoa trùng để sản xuất vaccine cho một làng nhỏ trong vùng dịch tễ. Một cách tiếp cận vaccine khác đó là nhận diện các epitope trong các giai đoạn khác nhau của Plasmodium đặc hiệu miễn dịch cho các tế bào T và B. Khi đã nhận diện được các epitope này thì có thể tạo ra được các vaccine peptit tổng hợp chứa các epitope này hoặc có thể tạo ra các kháng thể đơn clone đặc hiệu với các epitope này. Bằng cách này người ta đã phát hiện được kháng nguyên đích trong vaccine thoa trùng chiếu tia X đó chính là kháng nguyên CS. Kháng nguyên CS của P. falciparum chứa 412 acid amin có vùng trung tâm gồm khoảng 40 đoạn lặp lại với thứ tự các acid amin là Asn - Ala - AsnPro. Đoạn lặp lại này cấu tạo thành epitope dành cho tế bào B. Ngoài ra các kháng thể đơn clone đặc hiệu với những đoạn lặp lại này trên kháng nguyên CS có tác dụng bảo vệ chuột nhắt chống lại Plasmodium sống khi gây nhiễm. Điều này gợi ý rằng các vaccine peptit tổng hợp dựa trên đoạn lặp lại này có thể sinh ra các kháng thể bảo vệ. Để đưa ra loại vaccine peptit tổng hợp cần phải xác định số lượng các đoạn lặp lại hoạt động như epitope đặc hiệu cho tế bào B. Những thực nghiệm với những peptit tổng hợp gồm từ 1 - 5 đoạn lặp lại đã cho thấy peptit gồm 3 đoạn lặp lại là một epitope hoàn chỉnh, epitope này có thể phong bế hoàn toàn việc gắn của kháng thể vào thoa trùng. Trong một thử nghiệm, người ta đã gắn peptit tổng hợp gồm 3 đoạn lặp lại với giải độc tố uốn ván và dùng táchất là alum rồi tiêm bắp cho 35 người đàn ông khỏe mạnh tình nguyện (với liều 160 (g) thì thấy 71% trong số này đã sinh ra kháng thể kháng lại vaccine. 3 trong số người tình nguyện có nồng độ kháng thể cao và 4 người tình nguyện khác không dùng vaccine đã được thử thách bằng cách cho muỗi anophel nhiễm P. falciparum đốt. Kết quả cho thấy cả 4 trường hợp sau đều xuất hiện thể phân liệt (merozoit) trung bình sau 8,5 ngày, ngược lại 3 trường hợp trước thì có 1 người không thấy xuất hiện thể phân liệt hay bất cứ triệu chứng nào của sốt rét và 2 người còn lại có xuất hiện thể phân liệt nhưng mãi đến ngày thứ 11 mới xuất hiện, như vậy là muộn hơn so với nhóm chứng. Các nghiên cứu khác cũng cho kết quả tương tự. Các kết quả nghiên cứu mặc dù là có hứa hẹn như vậy nhưng cũng mới chỉ là bước đầu. Việc thử vaccine như trên cũng đã nêu lên một khó khăn chung của vaccine gồm có 1 peptide tổng hợp đóng vai trò là epitope đặc hiệu cho tế bào B được để kết hợp với 1 protein tải không liên quan là giải độc tố uốn ván đó là: mặc dù tạo ra được kháng thể chống peptit tổng hợp nhưng đáp ứng của tế bào T thì lại chống protein tải giải độc tố uốn ván. Vì vậy vaccine này không sinh ra được tế bào T có trí nhớ miễn dịch đặc hiệu với Plasmodium. Điều cần thiết là vaccine phải có cả epitope đặc hiệu cho tế bào B lẫn epitope đặc hiệu cho tế bào T. Người ta đã thử tìm epitope đặc hiệu cho tế bào B trong kháng nguyên CS bằng cách dùng một vaccine đậu bò tái tổ hợp có biểu hiện kháng nguyên CS để gây miễn dịch cho chuột thuần chủng. Thử nghiệm được tiến hành trên các dòng thuần chủng khác nhau nhưng chỉ có các dòng chuột nhắt mang các kháng nguyên hòa hợp mô lớp II là IAb và IAk mới có khả năng sản xuất nhiều kháng thể kháng kháng nguyên CS, điều này gợi ý rằng phân tử hòa hợp mô lớp II IAb và IAk đã trình iện các peptit của kháng nguyên CS cho các tế bào Th và bởi vậy gây ra sự hoạt hóa tế bào B. Để phát hiện các epitope đặc hiệu cho tế bào T trên kháng nguyên CS đã được nhận dạng bởi chuột có phân tử IAk người ta đã phân tích các đoạn peptit thẳng của kháng nguyên CS bằng chương trình máy tính để tìm ra các peptpit có các vòng xoắn (có biểu hiện amphipathic rõ rệt). Những peptit này thể hiện cả hai bề mặtái nước và kỵ nước, chúng được được giả thiết là gắn với các phân tử hòa hợp mô và các thụ thể tế bào T. Đoạn pepetit ký hiệu là Th2R có chỉ số amphipathic cao nhất. Người ta đãùng các peptit tổng hợp chứa epitope đặc hiệu cho tế bào B có chứa peptit Th2R để gây miễn dịch cho chuột IAk thì thấy chuột có đáp ứng kháng thể cao. Tiếp tục quá trình này người ta hy vọng sẽ tìm ra được các epitope đặc hiệu cho tế bào T và có thể chế ra được một vaccine hữu hiệu bằng cách kết hợp epitope đặc hiệu cho tế bào B với epitope đặc hiệu cho tế bào T. Kháng nguyên CS hình như có tính sinh miễn dịch thấp đối với tế bào T của người. Ví dụ khi nuôi Lympho bào máu ngoại vi của người sống trong vùng dịch tễ sốt rét với sự có mặt của các peptit tổng hợp gối lên nhau để có chiều dài bằng với phân tử kháng nguyên CS thì nhận thấy 40% số người này không có phản ứng tăng sinh lýmpho bào để đáp ứng với bất kỳ một peptit nào. Trong số các peptit gây phản ứng tăng sinh ở 60% mẫu tế bào có 2 peptit là peptit 20 (Th2R) và peptit 24 có chỉ số amphipathic cao nhất. Khi so sánh sự thay đổi thứ tự của các peptit CS người ta nhận thấy hai peptit trên có sự thay đổi trình tự các acid amin, vì vậy gây ra sự thay đổi kháng nguyên của ký sinh trùng sốt rét làm cho ký sinh trùng sốt rét có thể thoát khỏi đáp ứng miễn dịch. Vì vậy cần phải sàng lọc để tìm ra các các đoạn peptit ít có sự thay đổi. Sự biến đổi của các epitope đặc hiệu cho tế bào T nằm trong phân tử kháng nguyên CS là một giới hạn chính trong việc sinh ra đáp ứng miễn dịch qua trung gian tế bào chống ký sinh trùng sốt rét. Nhiều tài liệu đã chứng minh rằng đáp ứng miễn dịch qua trung gian tế bào hoạt động phối hợp với các kháng thể hoặc hoạt động một cách độc lập có vai trò quan trọng trong sốt rét. Chuột nhắt khi đã được gây miễn dịch bằng thoa trùng chiếu tia X thì có thể kháng lại sự nhiễm thoa trùng sống khi thử thách. Tuy nhiên, nếu sử dụng kháng thể kháng CD8 tiêm cho những chuột đã miễn dịch này thì khả năng miễn dịch chống thoa trùng bị mất đi. Ngoài ra những tế bào T phân lập từ những chuột đã miễn dịch này có thể gây ra được đáp ứng miễn dịch vay mượn trên những chuột khác. Đáp ứng miễn dịch qua trung gian tế bào chống sốt rét được thực hiện cả bằng hai kiểu: kiểu quá mẫn muộn và kiểu gây độc bởi tế bào Tc. Tế bào TCD4 có thể nhận biết các kháng nguyên thoa trùng đã kết hợp với phân tử hòa hợp mô lớp II ở trên bề mặt tế bào Kupffer. Đích tấn công của tế bào TCD8 + là những kháng nguyên thoa trùng được trình diện bởi các phân tử hòa hợp mô lớp I trên các tế bào gan bị nhiễm. Thoa trùng có thể thoát khỏi các đáp ứng miễn dịch qua trung gian tế bào và làm xuất hiện một lượng lớn thể phân liệt vì vậy làm giảm khả năng loại bỏ Plasmodium bằng hệ thống miễn dịch. Bảng dưới đây nói lên ảnh hưởng của việc loại bỏ các tế bào TCD4 hoặc TCD8 lên đáp ứng miễn dịch chống Plasmodium ở chuột đã được gây miễn dịch bằng vaccine thoa trùng chiếu tia X. Đáp ứng miễn dịch chống các bệnh giun sán Khác với các đơn bào, vi sinh vật chỉ có 1 tế bào và thường cư trú bên trong tế bào của người, giun sán là những vi sinh vật lớn gồm nhiều tế bào không cư trú bên trong tế bào và thường cũng không nhân lên ở trong cơ thể người. Mặc dù giun sán dễ tiếp cận với hệ thống miễn dịch của người hơn là những đơn bào và phần lớn các cá thể chỉ bị nhiễm ít loại giun sán. Vì lý do đó hệ thống miễn dịch cũng không đáp ứng mạnh và mức độ đáp ứng miễn dịch sinh ra cũng yếu. Các loài giun sán gây ra rất nhiều bệnh ở cả người và động vật. Hơn một tỷ người bị nhiễm giun đũa, một loại giun tròn cư trú ở ruột non; hơn 300 triệu người bị nhiễm Schistosoma, một loại sán sống trong máu gây ra trạng thái nhiễm mạn tính. Nhiều loại giun sán gây bệnh cho gia súc và xâm nhập vào người qua đường ăn uống đó là sán lợn, sánbò hoặc giun xoắn. KSTSR xâm nhập vào vật chủ, một loạt cơ chế bảo vệ xảy ra tạo nên tình trạng chống nhiễm bệnh, KSTSR luôn tìm cách xâm nhập vào tế bào vật chủ để sống và sinh sản, chúng phải đề phòng và tránh thoát hệ thống bảo vệ vật chủ và khi đó vật chủ cũng hình thành các cơ chế bảo vệ chống lại KSTSR rất phức tạp. Xét về mặt dịch tễ sốt rét, một vấn đề phức tạp, nhất là hình thái SR trong cộng đồng có thể thay đổi lớn từ vùng này đến vùng khác. Macdonald đã nêu 2 hình thái của dịch tễ sốt rét là ổn định và không ổn định. Tại vùng SR không ổn định tỉ lệ bệnh thường thấp, mức độ miễn dịch trong cộng đồng cũng thấp nhưng có thể sẽ xảy ra dịch trong một giai đoạn ngắn, bệnh ảnh hưởng tới mọi lứa tuổi. Ở vùng SR ổn định tỉ lệ bệnh cao, mức độ miễn dịch trong cộng đồng cao nên ảnh hưởng chủ yếu đến trẻ em (Wernsdorfer và cs., 1990). Những điều tra dịch tễ ở miền Tây châu Phi - nơi có sự lan truyền SR cao cho thấy tỉ lệ KSTSR trong cộng đồng là 50-60%, đặc biệt tỉ lệ nhiễm KSTSR ở trẻ em dưới 9 tuổi là 70-90%; tỉ lệ đáp ứng miễn dịch cao ở tất cả các lứa tuổi, phản ứng IFA dương tính 100% (Molineaux và cs., 1980). Hai vùng này, các nhà nghiên cứu thấy mức độ miễn dịch có liên quan đến triệu chứng lâm sàng và mật độ KSTSR. Lượng KSTSR giảm dần theo nhóm tuổi, ngược lại mức độ miễn dịch tăng theo tuổi. Ký sinh trùng sốt rét “lẩn tránh” hệ miễn dịch Các nhà khoa học Mỹ vừa phát hiện ra các ký sinh trùng sốt rét đã chơi trốn tìm với hệ miễn dịch của người. Ký sinh trùng sốt rét có tên khoa học Plasmodium falciparum (P.falciparum) đã liên tục thay đổi “diện mạo” của một protein mà nó đặt lên bề mặt của tế bào người như một thứ vũ khí tấn công. Tốc độ thay đổi của protein này quá nhanh khiến cho hệ miễn dịch không có đủ thời gian để tạo kháng thể chống lại nó. Trên thế giới hiện nay có ít nhất 300 triệu người mắc bệnh sốt rét, trong đó có ít nhất một triệu người chết mỗi năm, phần lớn tập trung ở các nước đang phát triển. Phát hiện mới đã gây bất ngờ về mức độ tiến hóa của ký sinh trùng P. falciparum, cho phép nó duy trì tuổi thọ lâu dài, từ lúc ký sinh trên muỗi cho đến khi tấn công cơ thể người. Bí quyết của P. falciparum chính là khả năng thay đổi liên tục phiên bản của một protein có tên là PfEMP1 - "quả mìn" mà nó gài trên bề mặt của các tế bào "con mồi". Tốc độ thay đổi protein PfEMP1 cực nhanh. Khi hệ miễn dịch còn đang lấn bấn tìm cách nhận diện để tạo kháng thể đối phó thì PfEMP1 đã "thay áo" trở thành một dạng khác. Cứ như thế, trò chơi trốn tìm lại được lặp lại. Các nhà khoa học đã biết được một họ gene có tên là var chuyên kiểm soát quá trình sản xuất protein PfEMP1. Trong hệ gene của ký sinh trùng sốt rét có ít nhất 50 gene var. Song các gene này không hoạt động đồng thời, mà chỉ có duy nhất một gene hoạt động vào từng thời điểm. Có thể nói, trong quá trình truyền bệnh của ký sinh trùng sốt rét, diện mạo của nó sẽ thay đổi từ một gene var này sang gene var khác, và trong khi một gene được bật lên thì tất cả các gene còn lại đồng loạt tắt. Nhóm nghiên cứu đến từ Viện y học Howard Hughes đã bắt tay tìm hiểu vì sao các gene var lại có khả năng thống nhất và hoạt động nhịp nhàng như vậy. Họ phát hiện ra nguyên nhân nằm ở ADN của các gene var, trong đó một protein đặc biệt được ví như "bộ máy điều chỉnh thông tin im lặng 2" - SIR2 - đóng vai trò làm "câm" ADN ở những gene var "im lặng". Theo tiến sĩ Alan Cowman, trưởng nhóm nghiên cứu, việc gỡ nhỏ các mắt xích trong cơ chế bật - tắt các gene var có thể mở ra các hướng mới trong việc phát triển thuốc chống sốt rét Ẩn mình vào trong tế bào hình cây của lách: Vẫn còn nhiều vấn đề chưa biết đến! Các chuyên gia đã khám phá cách ký sinh trùng sốt rét có thể tồn tại trong một loại bạch cầu trong lá lách, một bước đột phá có thể dẫn đến những loại thuốc và vắc xin phòng chống sốt rét mới.Thông thường, người ta tin sự phát triển của ký sinh trùng bị giới hạn ở các tế bào gan và hồng cầu.Bà Michelle Wykes, tác giả nghiên cứu thuộc Viện Nghiên cứu y khoa Queensland (Úc) cho biết, những phát hiện này mở ra một cách tiếp cận hoàn toàn mới để phát triển các loại thuốc và vắc xin. “Nghiên cứu của chúng tôi đã phát hiện ra cách bạch cầu ra lệnh tế bào tua hoạt động sai và che chắn cho các ký sinh trùng sốt rét khỏi sự tấn công hệ miễn dịch của cơ thể. Tế bào tua thường hoạt động giống tướng quân đội, ra lệnh cho các tế bào miễn dịch của cơ thể chống lại sự viêm nhiễm”, bà Wykes nói. “Hệ thống này thường hoạt động rất tuyệt vời. Tuy nhiên, vấn đề với bệnh sốt rét là nó đã tìm ra cách để ngăn chặn các tế bào tua thực hiện công việc của mình, nghĩa là căn bệnh khống chế phản ứng miễn dịch của chúng ta. Vì thế, người ta bị bệnh”, bà Wykes giải thích. Hơn 20 năm trước đây, bệnh sốt rét còn lưu hành rộng tại một số địa phương. Nếu tổ chức công tác giám sát, điều tra dịch tễ sốt rét ở cơ sở hoặc khi bệnh nhân sốt rét vào bệnh viện điều trị, một triệu chứng lâm sàng thường được phát hiện là lách sưng to, lá lách có thể sờ thấy được dưới hạ sườn trái với các mức độ khác nhau. Lách to là một triệu chứng đặc trưng của bệnh sốt rét. Bệnh nhân càng sống lâu trong vùng sốt rét lưu hành thì lách càng to. Vì vậy, lách to đã được sử dụng làm chỉ số đánh giá về mặt dịch tễ học sốt rét. Trong bệnh sốt rét, lách góp phần vào việc loại trừ các hồng cầu nhiễm ký sinh trùng một cách tích cực, kể cả các hồng cầu có kháng nguyên sốt rét hòa tan trên màng. Về vi thể, những biến đổi lách do bệnh sốt rét có thể được ghi nhận như tăng sinh các đại thực bào và tế bào nội mạch để làm nhiệm vụ thực bào các hồng cầu nhiễm ký sinh trùng; nhiều mao mạch bị tắc nghẽn bởi các hồng cầu nhiễm ký sinh trùng, tạo ra những ổ hoại tử nhỏ rải rác trong phần tủy lách. Các xoang lách ứ đầy máu. Ở các bệnh nhân sốt rét thường xuyên bị tái phát và tái nhiễm bệnh thì sự tăng sinh tổ chức liên kết rất rõ rệt làm cho mức độ của lách chắc hơn. Ở một số bệnh nhân sốt rét thấy xuất hiện hội chứng cường lách, những bệnh nhân này mặc dù cơ thể sạch ký sinh trùng trong máu từ lâu nhưng hồng cầu, có khi cả bạch cầu và tiểu cầu đều giảm thấp. Trường hợp lách to ở người mới sốt, khi hết sốt, hết ký sinh trùng sốt rét thì lách có thể co nhỏ lại. Những người sống trong vùng sốt rét lưu hành có thể có lách to, có kháng thể sốt rét cao, gan to… Vì vậy, có quan niệm cho rằng đó là hội chứng lách to nhiệt đới (tropical splenomegaly syndrom); cũng có quan niệm cho rằng đó là sự đáp ứng miễn dịch quá mức đối với sốt rét. Có thể nói rằng lách có chức năng quan trọng giúp bảo vệ, chống lại bệnh sốt rét vì nó có khả năng giữ lại hồng cầu nhiễm ký sinh trùng và cả hồng cầu bình thường có sự nhạy cảm với kháng thể sốt rét hòa tan. Lách còn tham gia khá sớm vào cơ chế miễn dịch tế bào và dịch thể nhờ có đại thực bào và những kháng thể được sản sinh trong lách. Vì vậy, ngay sau khi ký sinh trùng đột nhập vào cơ thể người, lách có thể tăng thể tích do sung huyết. Trong sốt rét thể thông thường, lách to ít, chưa sờ thấy lúc đầu; ít tuần sau, lách to vừa, mềm và có màu đỏ tím. Trường hợp sốt rét tái nhiễm, lách to tương đối nhanh, trọng lượng có thể từ 500g đến 2-3kg, chắc, có màu xám. Đối với sốt rét thể ác tính, lách to, có màu đỏ thẫm, lấm tấm các ổ chảy máu nhỏ. Người bình thường, bờ dưới của lách không sờ thấy được dưới hạ sườn trái. Khi bị mắc bệnh sốt rét, lách thường sưng to lên nhưng không phải bất cứ bệnh nhân sốt rét nào cũng có lách to. Lách chỉ to khi bị nhiễm ký sinh trùng sốt rét nhiều lần và không được điều trị đúng. Theo quy định, lách sưng to được chia làm 4 mức độ từ nhỏ đến lớn. Độ 1 có bờ dưới lách đến gần 1/4 đường từ mạng sườn trái tới rốn. Độ 2 có bờ dưới lách nằm ở 1/4 đến 1/2 đường từ mạng sườn trái tới rốn. Độ 3 có bờ dưới lách nằm quá 1/2 đường từ mạng sườn trái tới rốn. Độ 4 có bờ dưới lách ngang hoặc quá rốn. Khi khám lâm sàng, cần phải xác định lách to do nguyên nhân nhiễm ký sinh trùng sốt rét hay lách to do những nguyên nhân bệnh lý khác. Khi mới bị nhiễm bệnh sốt rét, có khoảng 60-70% bệnh nhân lách sưng to, hơi đau, sờ thấy dưới bờ sườn trái, chưa rắn chắc, thường ở độ 1 và độ 2; lách có thể trở về mức bình thường nếu được điều trị tốt. Nếu bị mắc bệnh sốt rét tái diễn nhiều năm, trải qua các đợt tái phát rồi tái nhiễm liên tiếp nhau mà không được điều trị tốt, nhất là những người dân sống trong vùng sốt rét lưu hành nặng thì lách sẽ bị xơ hóa, rắn chắc và khả năng không co lại được. Lách to dễ bị giập vỡ. Biến chứng giập vỡ lách thường gặp ở những bệnh nhân mới mắc sốt rét, hiếm gặp hơn ở những người mắc sốt rét đã lâu. Sở dĩ như vậy vì khi mới mắc sốt rét, lách to nhanh nên yếu và dễ vỡ. Tuy nhiên, sự giập vỡ lách còn tùy thuộc vào cơ địa của từng bệnh nhân. Khi lách to do bị mắc bệnh sốt rét, sự tiến triển có hai khả năng xảy ra: lách không thể trở lại mức độ bình thường như cũ dù rằng bệnh nhân đã khỏi hẳn bệnh sốt rét vì các tế bào của lách đã bị kết xơ, mao mạch, thần kinh giao cảm và phó giao cảm không thể phục hồi lại được hoặc lách có thể trở lại bình thường nếu bệnh nhân được điều trị tốt và không bị phát, tái nhiễm lại bệnh sốt rét. Một số trường hợp lách quá to ảnh hưởng đến sinh hoạt và khả năng lao động. Lách to và ý nghĩa về mặt dịch tễ học sốt rét Lách to phản ánh mức độ bệnh sốt rét lưu hành ở một địa phương và mức độ miễn dịch trong nhân dân. Với một cộng đồng dân cư mới đến vùng sốt rét lưu hành, người dân dễ có nhiều nguy cơ bị mắc bệnh sốt rét; khi tổ chức điều tra, giám sát dịch tễ ghi nhận tỷ lệ nhiễm ký sinh trùng sốt rét cao hơn tỷ lệ lách to. Với người dân sống lâu năm trong vùng sốt rét lưu hành, tỷ lệ lách to có thể cao hơn tỷ lệ nhiễm ký sinh trùng. Mức độ của lách to cũng phụ thuộc vào chủng loại ký sinh trùng sốt rét bị nhiễm. Nhiễm ký sinh trùng sốt rét Plasmodium vivax thường thấy lách sưng to, nhiễm ký sinh trùng sốt rét Plasmodium falciparum lách sưng trung bình, nhiễm ký sinh trùng sốt rét Plasmodium malariae ít gây sưng lách. Nhờ các biện pháp can thiệp có hiệu quả, bệnh sốt rét tại các vùng sốt rét lưu hành của nước ta đã có những chuyển biến tốt. Hiện nay, bệnh nhân sốt rét vào điều trị ở bệnh viện ít phát hiện được triệu chứng lách sưng to do bệnh sốt rét. Vấn đề này đã khẳng định mức độ lưu hành bệnh ở các vùng sốt rét không còn nghiêm trọng như trước đây và sự miễn dịch đối với bệnh trong cộng đồng người dân sẽ giảm dần. Tuy nhiên, nguy cơ dịch sốt rét xảy ra là điều mà các cơ sở y tế luôn luôn cần phải cảnh báo. Miễn dịch dịch thể đặc hiệu từng giai đoạn phát triển KSTSR TSR phát triển trong cơ thể qua các giai đoạn: thoa trùng xâm nhập cơ thể người, KST phát triển ở tế bào gan, KST phát triển ở máu ngoại vi và qua đó cơ thể hình thành các đáp ứng miễn dịch như sau: -Miễn dịch giảm thoa trùng: qua nghiên cứu thực nghiệm trên những người tình nguyện người ta thấy có sự liên quan giữa kháng thể (KT) bảo vệ và KT phản ứng với KN bề mặt của thoa trùng.Miễn dịch bảo vệ chống thoi trùng do KT đặc hiệu ngăn cản thoa trùng xâm nhập vào tế bào gan và có thể làm sạch thoa trùng ở máu ngoại vi. Sự phát hiện KT đặc hiệu chống thoa trùng là một thông tin quan trọng cho cuộc nghiên cứu vaccine chống SR trong thời gian tới. -Miễn dịch của thể ngoài hồng cầu: khi KST xâm nhập vào tế bào gan thì KN là thoa trùng, là các thể của KSTSR phát triển ở tế bào. Đáp ứng miễn dịch chưa thấy biểu hiện rõ, KT chống lại KSTSR ở gan chưa được nghiên cứu chi tiết. -Miễn dịch thể vô tính ở hồng cầu: Nghiên cứu đáp ứng MD của cơ thể đối với KSTSR ở giai đoạn hồng cầu được nghiên cứu cho thấy KT có vai trò trực tiếp ức chế merozoites xâm nhập vào hồng cầu, làm trung hoà độc tố và hoạt hoá bổ thể nên nó làm tăng hiện tượng thực bào. Khi KST sống và phát triển trong hồng cầu, miễn dịch qua trung gian tế bào (CMI- Cellular Mediated Immune) được hình thành, lympho T hoạt hoá sản xuất các lymphokin giúp đại thực bào giết KST và nuốt các hồng cầu bị nhiễm. -Miễn dịch đối với thể giao bào: Thoa trùng xâm nhập vào tế bào gan, KST phát triển trong tế bào gan và hồng cầu. Sau một thời gian ngắn ở máu ngoại vi sẽ xuất hiện giao bào và tuỳ loại KSTSR (P.falciparum sau 9-10 ngày, P.vivax sau 2-3 ngày). Trong huyết thanh những người có miễn dịch thường có kháng thể phản ứng chéo với cả hai thể vô tính và giao bào. Ngoài ra, các nghiên cứu gần đây cho biết KT đặc hiệu với giao bào trong vùng SRLH cao. Điều này giúp các nhà khoa học nghiên cứu vaccine loại chống giao bào. Những năm gần đây, nhiều tác giả đã ứng dụng các kỹ thuật miễn dịch trong nghiên cứu dịch tễ và giám sát hiệu quả các biện pháp phòng chống SR (Wernsdorfer, 1990). Theo dõi nhóm trẻ được bảo vệ và không được bảo vệ ở miền Tây châu Phi thấy có đáp ứng KT (IFA dương tính) ở nhóm được bảo vệ giảm dần từ 100% (năm đầu tiên) xuống còn 65% (năm thứ 2) và tiếp tục giảm vào những năm sau. Nhóm không được bảo vệ không có sự thay đổi, 100% IFA dương tính ở các năm (Molineaux và Gramicia, 1980). Những nghiên cứu về P. falciparum đã phát hiện được thêm về tính đa dạng di truyền của những mẫu phân lập thu được tại nhiều vùng địa lý khác nhau trên thế giới và ngay cả trong một quốc gia nhưng nơi có sinh cảnh khác nhau. Tính đa dạng này dàn rộng ra trên 6 biến thể enzyme như gluco-phosphate isomerase (GPI), 6-phosphate gluconate dehydrogenase (PGD), lactate dehydrogenase (LDH), adenosine deaminase (ADA) và peptidase E (PEPE), trên sự sắp xếp trình từ ADN,trên các dạng proteine khác nhau, trên các quyết định KN và trên các mức độ thụ cảm của KST với thuốc SR. Mức độ đa dạng KN cũng liên quan đến cường độ truyền bệnh của từng vùng riêng biệt vàchính sự đa dạng về KN này dẫn đến một số kết quả trái ngược, không đồng nhất với phương pháp nhuộm giêm sa của các RDTs. Cho muỗi đốt để miễn dịch với bệnh sốt rét - Nên chăng? Các nhà khoa học châu Âu đã thử nghiệm việc sử dụng muỗi để đưa vắcxin chống bệnh sốt rét vàng da vào cơ thể người tình nguyện. Kết quả thử nghiệm đã thành công ngoài dự kiến. Những người được tiêm thuốc bằng cách cho muỗi đốt đều miễn dịch với căn bệnh sốt rét này. Tuy thí nghiệm không được tiến hành rộng rãi nhưng dù sao thành công này cũng cho thấy các nhà khoa học có thể đang đi đúng hướng trong việc tìm ra một loại vắcxin chống lại căn bệnh sốt rét nguy hiểm. Bác sĩ Carlos Campbell, làm việc cho PATH (Chương tình sử dụng những công nghệ thích hợp cho sức khỏe) ở Seattle, Mỹ, cho rằng: "Vaccine cho bệnh nhân sốt rét đang đi từ phòng thí nghiệm vào cuộc sống". Từ trước đến nay, chế tạo vắcxin phòng sốt rét vàng da là một quá trình rất khó khăn. Thuốc đặc trị Chloroquine chỉ có thể giết ký sinh trùng gây bệnh khi chúng đã vào tới máu, mà đây đã là giai đoạn nguy hiểm nhất với bệnh nhân. Thí nghiệm này cho thấy, trong tương lai không xa, người ta có thể dùng kí sinh trùng hoàn chỉnh của bệnh sốt rét trong việc chế tạo vắcxin phòng bệnh. Hàng năm trên thế giới, có khoảng 1 triệu người tử vong vì sốt rét vàng da, trong đó đa số là trẻ em dưới 5 tuổi ở châu Phi.
|