|
Entamoeba histolytica: Tác nhân ký sinh trùng gây tử vong đứng thứ 2 trên thế giới? (Phần 4-còn nữa)
Tiếp theo Phần 3
E. histolytica xâm nhập, phá hủy lớp niêm nhầy ruột: Vai trò enzyme protease cysteine A5
Các enzyme protease cysteine (CP) đóng vai trò then chốt trong quá trình gây bệnh của E. histolytica. Trong đó, protease cysteine A5 (CP-A5, EHI_168240) chỉ có ở loài E. histolytica, đây là yếu tố quan trọng của amíp giúp loại bỏ chất nhầy vì nó nhắm vào peptide ở vùng có ít sự glycosyl hóa của MUC2.Enzyme này chỉ xuất hiện bề mặt của thể tư dưỡng hoạt động có tính liên kết integrin Arg-Gly-Asp (RGD) cũng được mô tả ở cathepsin Xnhân thực bậc cao. CP-A5 liên kết thông qua motif RGD với integrin αVβ3 trên các tế bào giống tế bào ruột và kích thích phản ứng viêm. Tương tác của CP-A5 amíp với integrin αVβ3 kích hoạt tín hiệu bên ngoài-bên trong các tế bào hình đài dẫn đến tiết mucin cùng với sự kích hoạt của một số kinase (kinase thuộc họ SRC, kinase bám dính tập trung FAK, PI3K, PKCδ và MARCKS). Một enzyme giống cyclooxygenase của E. histolytica điều chỉnh hoạt động của CPA5 thông qua tương tác protein-protein giúp ổn định CP-A5 và tăng độc lực. Dữ liệu gần đây cho thấy vai trò CP-A5 trong việc phân hủy màng sinh học của vi khuẩn. Để xâm nhập vào mô, thể hoạt động làm rối loạn cấu trúc giàn “scaffold collagen” lỏng lẻo để trốn thoát vào lớp niêm mạc. Thể hoạt động bị bất hoạt biểu sinh đối với CP-A5, không có hoạt tính proteinase của nó, không gây ra phản ứng viêm mô hoặc không kích hoạt quá trình tái cấu trúc collagen cần thiết để xâm nhập lớp đệm niêm mạc (lamina propria). Trong mô hình đại tràng mô cấy, các hoạt động enzym của matrix metaloproteinase của người (MMP-1 và -3) được xác định là chịu trách nhiệm về thay đổi cấu trúc sợi collagen trong quá trình xâm nhập đường ruột bởi E. histolytica. Rõ ràng, hoạt động CP-A5 chuyển pro-MMP-3 thành enzyme MMP-3 hoạt động, sau đó lại kích hoạt pro-MMP-1, dẫn đến phân huỷ collagen. Ngoài vai trò trong bệnh lỵ amíp đường ruột, hoạt động của CP-A5 cũng cần thiết để gây ra sự hình thành các tổn thương ở gan ở chuột hamstervà liên quan đến sự mất đi khung tế bào actin và tổ chức của cấu trúc gắn kết tập trung gây ra chết tế bào nội mô. Nhìn chung, CP-A5 là một enzyme đa chức năng đóng vai trò quan trọng trong quá trình sinh bệnh của E. histolytica ở đại tràng bằng cách phân hủy chất nhầy và màng sinh học vi khuẩn, liên kết với thụ thể integrin αvβ3 trên các tế bào hình đài để gây ra phản ứng tiền viêm, kích hoạt quá trình tái cấu trúc cấu trúc collagen dạng sợi của ruột và còn đóng vai trò trong việc hình thành áp-xe gan. Tác động qua lại giữa E. histolytica gây bệnh và ruột người ở giai đoạn đầu bệnh amíp Để thành công trong quá trình xâm nhập đường ruột, E. histolytica phải có khả năng di chuyển và khung tế bào giàu actin đóng vai trò trung tâm trong quá trình xâm lấn phức tạp. Trong giai đoạn đầu quá trình xâm nhập đường ruột, sau khi lớp màng nhầy bảo vệ bị phá hủy, thể hoạt động gây hại thâm nhập vào nhu mô ruột, bám vào và phá hủy ma trận ngoại bào (ECM) thông qua hoạt động của các thụ thể bề mặt amíp nhận diện các thành phần ECM và hoạt động của proteinase. Thể hoạt động phát triển trong biểu mô đại tràng bằng cách gắn vào tế bào động vật có vú nhờ các thành phần glycosyl hóa bề mặt như lectin Gal-GalNAc, lipopeptidophosphoglycan (LPPG) gắn với glycocalyx, hoặc bằng cách tiếp xúc trực tiếp amíp-tế bào có sự góp mặt của các protein như KERP1, KERP2. Các protein giàu serine, threonine và isoleucine (STIRPs)và adhesin ADH112. Các proteinase phá vỡ mối nối tế bào-tế bào của các tế bào biểu mô, tiêu diệt tế bào động vật có vú và xảy ra hiện tượng thực bào. Ngoài ra, sau khi bám vào tế bào chủ, amíp có thể "cắn đứt" và ăn vào các mảnh vỡ tế bào chủ bằng cách nhấm nháp tế bào (trogocytosis). Phản ứng miễn dịch vật chủ bắt đầu bằng sự thâm nhập của bạch cầu trung tính vào các vị trí ký sinh trùng xâm nhập, tiếp theo là sự kích hoạt các tế bào lớn (mast cell), đại thực bào, tế bào T tiêu diệt tự nhiên và sau đó là tiết các cytokine tiền viêm. Bằng cách nghiên cứu phản ứng tiền viêm và tiết chất nhầy do E. histolytica gây ra, hai dữ liệu gần đây cho thấy: (i) Phản ứng tiền viêm tương quan với việc giảm điều hòa yếu tố phiên mã liên quan đến biệt hóa tế bào Math1 (atonal 1 chuột đồng đẳng), việc ức chế yếu tố này dẫn đến giảm số lượng tế bào hình đài trong quá trình phát triển mô; (ii) Hệ vi sinh vật đường ruột phục hồi các phản ứng bình thường đối với tiết chất nhầy và kích hoạt cytokine tiền viêm, trong mô hình chuột bị rối loạn hệ khuẩn ruột do dùng kháng sinh và nhiễm E. histolytica. 
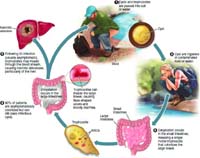
Chu kỳ sinh học về quá trình nhiễm Entamoeba histolytica
Những kết quả này trước tiên gợi ý rằng thể hoạt động có tác động trực tiếp đến việc điều hòa sự tồn tại của tế bào hình đài và thứ hai, vai trò của hệ vi sinh vật trong việc kiểm soát các phản ứng tiền viêm, có tác dụng bổ sung cho vai trò dinh dưỡng và bảo vệ choamíp được đề cập ở trên. Nghiên cứu về bản chất, hoạt động và tầm quan trọng (về khả năng gây bệnh) của "các yếu tố độc lực" amíp và tín hiệu hỗ trợ quá trình xâm nhập đường ruột đã được nghiên cứu chuyên sâu. Sự phá vỡ các mối nối giữa các tế bào biểu mô bởi E. histolytica Màng biểu mô hướng ra lòng ruột được cấu tạo bởi một lớp các tế bào phân cực (tế bào ruột, tế bào hình đài, tế bào nội tiết ruột (enteroendocrine), Paneth, tế bào M (microfold), tế bào cup và tế bào cảm ứng hóa chất) tạo thành hàng rào vật lý - hóa học và miễn dịch chống lại các kháng nguyên lòng ruột và hệ vi sinh vật đường ruột, đồng thời cho phép hấp thụ các chất dinh dưỡng và nước. Để gắn kết các tế bào với nhau cần có các mối nối chặt (Tight Junctions - TJs), mối nối dính (Adherens Junctions-AJs) và tiểu thể nối giãn bào (desmosome - DSMs). Đặc biệt, TJs điều chỉnh quá trình vận chuyển giữa các tế bào của các đại phân tử và ion qua lớp biểu mô. Các protein màng tích hợp của họ đặc hiệu TJ bao gồm occludin, claudin và phân tử kết nối (Junctional Adhesion Molecules-JAM) có trách nhiệm duy trì hàng rào ion của khoảng trống giữa các tế bào; claudin tương tác với các protein giàn scaffold như zona occludens (ZO-1 và ZO-2), liên kết chúng với protein nội bào khác và vi sợi actin. Occludin điều chỉnh dòng chảy của các đại phân tử, trong khi claudin kiểm soát dòng ion và hạn chế sự khuếch tán của protein và lipid, hoạt động như hàng rào màng và cùng với JAM, điều chỉnh tính phân cực của biểu mô. Proteinase cysteine CP112 từ loài E. histolytica đóng vai trò trong quá trình phân hủy các mối nối tế bào-tế bào. CP112 được mô tả với một vùng xúc tác chuẩn và một motif RGD, enzyme này phân hủy collagen type I, fibronectin và hemoglobin. Proteinase CP112, cùng với yếu tố bám dính ADH, tạo thành phức hợp độc lực EhCPADH. Thể hoạt độngbiểu hiện quá mức CP112 hoặc enzyme được tinh lọc gây ra giảm điện trở xuyên biểu mô của tế bào và phá vỡ chức năng hàng rào của các TJ ở tế bào giống tế bào ruột (tế bào Caco-2). Ngược lại, CP112 không ảnh hưởng đến độ thẩm thấu giữa các tế bào của các đại phân tử. Bằng cách sử dụng kính hiển vi đồng tiêu cự độ phân giải cao và xét nghiệm miễn dịch kết tủađã chứng minh rằng CP112 tinh khiết ảnh hưởng đến tính toàn vẹn của claudin-1 và claudin-2, không tìm thấy tác dụng trên claudin-4, occludin, ZO-1 hoặc ZO-2. Điều trị cho chuột bằng protease CP112 đã xác nhận những dữ liệu này, nghiên cứu bổ sung cho thấy thể hoạt động cũng có thể ảnh hưởng đến AJs và DSMs và chỉ ra vai trò của CP112 trong sự kiện độc lực nhiều yếu tố của E. histolytica. CP112 không phải là yếu tố duy nhất phá vỡ các TJ. Ví dụthể hoạt động tiết ra phân tử gây viêm prostaglandin E2 (PGE2), chất này kích thích tiết chloride trong các tế bào động vật có vú và phá vỡ hàng rào ion của các TJ bằng cách tác động lên claudin với tính thẩm thấu Na+ tăng, PGE2 từ E. histolyticatách claudin-4 khỏi màng tế bào động vật có vú ảnh hưởng đến hàng rào ion của TJs. Các protease bổ sung khác tham gia vào quá trình biến đổi protein TJ: CP-A5 (được thử nghiệm bằng nhiễm trùng amíp ở chuột Muc2−/−) và các protease serine tham gia vào việc giảm điện trở xuyên biểu mô của các tế bào biểu mô. 
Đường lây nhiễm và vị trí vùng thương tổn do amip Entamoeba histolytica
Do chức năng chính của mối nối tế bào là điều hòa vận chuyển ion trong ruột, việc phối hợp các hoạt động được mô tả ở trên sẽ là một điều thú vị, gồm cả sự phá vỡ các mối nối chặt bởi protease amíp, với hai bộ dữ liệu quan trọng liên quan đến giai đoạn đầu của nhiễm amíp. Dựa trên đặc tính gây độc tế bào của E. histolytica dẫn đến làm chết tế bào ở động vật có vú, các tác giả đã sàng lọc một thư viện RNA can thiệp của người (RNAi) để chọn các tế bào giống tế bào ruột có khả năng chống lại cái chết khi có mặt thể hoạt động gây hại. Việc xác định các gen được làm giàu rõ rệt, việc tắt chúng bằng RNAi sẽ làm tăng khả năng sống tế bào khi tiếp xúc với E. histolytica, làm nổi bật các kênh vận chuyển ion cho Ca2 ± phụ thuộc K+ (5 gen); Cl-(4 gen), Na+ (3 gen) và Ca2+ (2 gen). Sự ức chế thuốc đối với dòng ra K+ (K+ efflux) đã ngăn chặn sự chết đi của các tế bào biểu mô ruột và đại thực bào, cho thấy sự kích hoạt trực tiếp của dòng ra K+ như một dấu hiệu sớm của phản ứng mô sau khi E. histolytica xâm nhập đường ruột. Bộ dữ liệu thứ hai tương ứng với phân tích hệ phiên mã (transcriptome) và hệ protein (proteome) của các yếu tố của con người được điều chỉnh đáng kể ở giai đoạn đầu của quá trình tương tác giữa E. histolyticagây bệnh và mô hình giàn scaffold 3D của đại tràng. Các gen được điều hoà tăng cho các thụ thể hoạt độngnhư kênh ion đã được xác định: Tiểu đơn vị epsilon của thụ thể axit gamma-aminobutyric; kênh CL liên quan đến viêm loét đại tràng và tiểu đơn vị epsilon của thụ thể acetylcholine có thể mở các kênh ion. RNA trái ngược không mã hóa dài SLC9A3 1nhắm vào gen SLC9A3 mã hóa bộ trao đổi Na+/ H+ NHE3 liên quan đến tiêu chảy bẩm sinh cũng được điều chỉnh tăng. Ở cấp độ protein, các thành phần của thể liên kiết (desmosome) và các yếu tố liên kết chúng với khung tế bào (desmoplakin, plakophilin-3 và plectin-1) được tiết ra dồi dào; chúng quan trọng cho việc duy trì tính toàn vẹn của mô. Dữ liệu này cho thấy sự phá vỡ cấu trúc chịu trách nhiệm cho mối nối tế bào kết hợp với sự phá vỡ các kênh ion, là những tổn thương chính xảy ra ở giai đoạn đầu tiếp xúc giữa E. histolytica và biểu mô ruột, cả hai đều dẫn đến sự sụp đổ của cấu trúc mô. Do đó, hoạt động tiết chất nhầy, rò rỉ mô và bong tróc biểu mô là nguyên nhân gây ra các triệu chứng ban đầu của bệnh lỵ amíp. Chúng là giả thuyết cho cái chết được dự đoán của tế bào vật chủ, dù cách các tế bào biểu mô chết do tương tác với E. histolytica vẫn là một câu hỏi chưa có lời giải. 
Tổng quan về bộ gen E. histolytica HM-1:IMSS Clone 6-2001.
Biểu đồ này chỉ ra tòa bộ cấu trúc với chiều dài, chú giải và PacBio và Illumina đọc bộ gen của E. histolytica HM-1:IMSS Clone 6 2001. Từ ngoài vào trong, khung va chiều dài mới của chúng (block màu xám với một 1:10 kb), các đoạn trình tự liên tục thu được từ các đoạn reads (đoạn trình tự nhỏ) gối nhau của bộ gen trước đây (mã cột màu tím, trình tự mã hóa + sợi (mã cột màu xanh lá), trình tự mã hóa -sợi (mã cột màu cam), gen tRNA (mã cột màu đen), gen đơn bản sao (mã cột màu cam), độ bao phủ PacBio (một đường màu đen) và độ bao phủ Illumina (một đường màu xanh da trời) |
Sự chết tế bào động vật có vú khi tiếp xúc với Entamoeba histolytica Sau khi E. histolytica xâm nhập vào niêm mạc ruột, các tế bào sẽ bị chết và cho thấy sự phá hủy các tế bào động vật có vú yêu cầu tiếp xúc với thể hoạt động, kích hoạt dòng calci đi vào. Sự kết dính của thể hoạt động với tế bào ruột xảy ra thông qua các yếu tố khác nhau trên bề mặt amíp, gồm LPPG, lectin Gal-GalNAc, KERP1, STIRP và phức hợp CPAHD112. Sự tiếp xúc này dẫn đến chết tế bào người với tỷ lệ lớn, khoảng 40% tế bào giống tế bào ruột được nuôi cấy bị tiêu diệt trong vòng 30 phút sau khi tương tác. Các tế bào chết có thể được ăn vào bằng sự thực bào, đây là một quá trình quan trọng để loại bỏ mầm bệnh, tế bào chết hoại tử hoặc chết theo chương trình. Ở E. histolytica, thực bào là cách để lấy chất dinh dưỡng vì ký sinh trùng này là một tế bào thực bào chuyên nghiệp, ăn vi khuẩnvà các loại tế bào động vật có vú khác nhau. Tuy nhiên, sự tương tác giữa tế bào sống (biểu mô, hồng cầu hoặc lympho bào) và E. histolyticacó sự liên quan đến các bước phức tạp. Amíp khi gặp các tế bào động vật có vú sẽ nhanh chóng kích hoạt quá trình “nhấm nháp” tế bào (trogocytosis), một hiện tượng mà trong đó amíp cắn và ăn vào các mảnh vỡ tế bào vật chủ riêng biệt, góp phần tiêu diệt tế bào. Việc hấp thu và hiển thị các protein màng của tế bào động vật có vú trên bề mặt của amíp bảo vệ ký sinh trùng khỏi bị ly giải bằng cách ức chế chuỗi phản ứng bổ thể, cho thấy sự “nhấm nháp” tế bào góp phần vào các chiến lược phòng thủ của amíp để trốn tránh miễn dịch. Sự nhấm nháp tế bào và thực bào xảy ra bằng các cơ chế khác nhau, dù nhiều bước phân tử ban đầu có thể giống nhau. 
Dạng tư dưỡng của các amip khác nhau

Dạng bào nang amip của các vi sinh vật khác nhau
Giết chết tế bào phụ thuộc vào tiếp xúc bởi E. histolytica kích hoạt quá trình chết theo chương trìnhtế bào T và bạch cầu trung tính thông qua kích hoạt protein caspase 3 của tế bào vật chủ, nhưng các cơ chế chết không phụ thuộc caspase ở tế bào T cũng đã được mô tả, chúng bao gồm khử phosphoryl hoá tyrosine protein và tạo ra ROS và liên quan đến các kinase amíp, PI3K và PKC. Các dòng tế bào biểu mô ruột của người có thể trải qua quá trình chết theo chương trình (liên quan đến hoạt hóa caspase-3 và -9) khi tương tác với E. histolytica. Để tìm kiếm các cơ chế phân tử gây ra quá trình chết theo chương trình của tế bào ruột, các tác giả đã nghiên cứu các RNA không mã hóa, chẳng hạn như microRNA (miRNA), là các phân tử điều hòa âm của biểu hiện gen. Họ đã tiết lộ một tập hợp gồm sáu miRNA với các mạng đồng điều hòa miRNA-mRNA phức tạp, trong đó các miRNA nhắm vào các gen chống chết theo chương trình XIAP, API5, BCL2 và AKT1. Một trong số đó, miR-643 thực hiện điều hòa giảm sau phiên mã của XIAP bằng cách nhắm vào mRNA 3’-UTR của nó. Việc sử dụng các tác nhân đối kháng miR-643 dẫn đến việc phục hồi mức độ XIAP và ức chế quá trình chết theo chương trình và hoạt hóa caspase 3 và 9. Vai trò chức năng của trục miR-643 / XIAP trong quá trình chết theo chương trình của tế bào ruột được kích hoạt bởi E. histolytica. Vai trò các miRNA khác được phát hiện trong nghiên cứu này vẫn đang được tiếp tục thử nghiệm, cũng như vai trò của các RNA dài không mã hóa được điều hòa tăng trong mô hình giàn scaffold 3D của đại tràng người. Các LncRNA tham gia vào nhiều quá trình của tế bào và có thể tương tác với các miRNA để điều hòa vai trò của chúng, kể cả các sửa đổi biểu sinh điều hòa biểu hiện gen. Lĩnh vực nghiên cứu này còn là điều bí ẩn đối với nhiễm amíp và tiến triển của bệnh lỵ amíp. Tuy nhiên, những tiến bộ thú vị cũng đã gặt hái được trong nghiên cứu về tương tác giữa ký sinh trùng đường ruột với vi khuẩn. Cảm ứng miRNA có tương quan với độ phong phú tương đối của một số phân loại vi sinh vật, cho thấy các vi sinh vật điều hòa cảm ứng miRNA chủ yếu thông qua các chất chuyển hóa. Ngược lại, các miRNA do tế bào biểu mô đường ruột tiết ra làm thay đổi thành phần vi sinh vật. Chắc chắn rằng, những hướng nghiên cứu mới liên quan đến việc kiểm soát tác dụng gây độc tế bào của amíp bởi các tế bào của người kêu gọi các nghiên cứu xác định sự tham gia của RNA không mã hóa của người trong việc điều hòa sự chết tế bào ruột; đổi lại, cũng cần hiểu rõ vai trò của các RNA trái ngược tự nhiên đặc hiệu của amíp xuất hiện trong quá trình xâm lấn đại tràng của người trong việc kiểm soát các yếu tố độc lực.
(còn nữa) -->Tiếp theo Phần 5
|